The logic of consciousness. Part 2. Dendritic waves
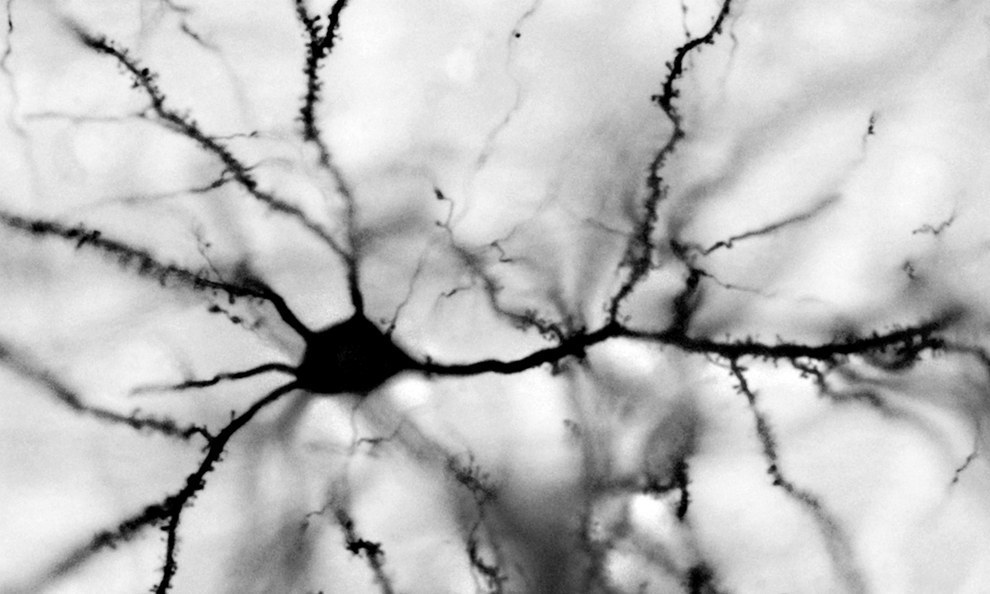 In the previous part, we showed that waves having a specific internal pattern can occur in a cellular automaton. Such waves can be launched from anywhere in the cellular automaton and propagate throughout the space of the automaton's cells, transferring information. It is tempting to suggest that the real brain can use similar principles. To understand the possibility of analogy, we’ll take a look at how the neurons of the real brain work.
In the previous part, we showed that waves having a specific internal pattern can occur in a cellular automaton. Such waves can be launched from anywhere in the cellular automaton and propagate throughout the space of the automaton's cells, transferring information. It is tempting to suggest that the real brain can use similar principles. To understand the possibility of analogy, we’ll take a look at how the neurons of the real brain work. The brain consists of gray and white matter. Gray matter is a brain structure made up of neurons and glial cells. White matter is the axons of neurons, they are nerve fibers. These fibers form the bonds of some brain structures with others.
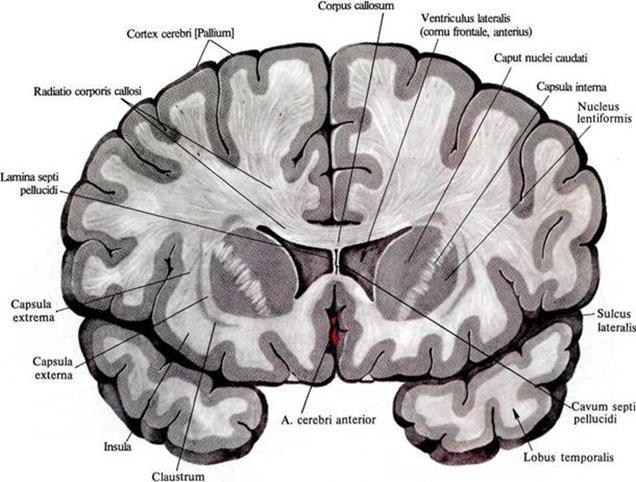
Distribution of white and gray matter in the frontal section of the brain
Structures that are closer to the center of the brain are usually referred to the ancient brain. The ancient brain unites us with animals and implements mechanisms honed by evolution and more or less common to many living things. The bulk of human gray matter is in the cortex. The cortex is a layer of gray matter with a thickness of 1.3 to 4.5 mm that makes up the outer surface of the brain. There are many arguments in favor of the fact that the cortex, unlike the ancient brain, does not implement genetically based algorithms, but is capable of learning and self-organization.
The main brain cells are neurons and glial cells. Both appear to play a significant role in information processes. To simplify the story, for now, we will only talk about neurons. Talk about glial cells is postponed for some time.
Neurons come in many types. The most massive neurons in the cortex are pyramidal neurons. They account for 75% of all neurons in the cortex. The figure below shows them.
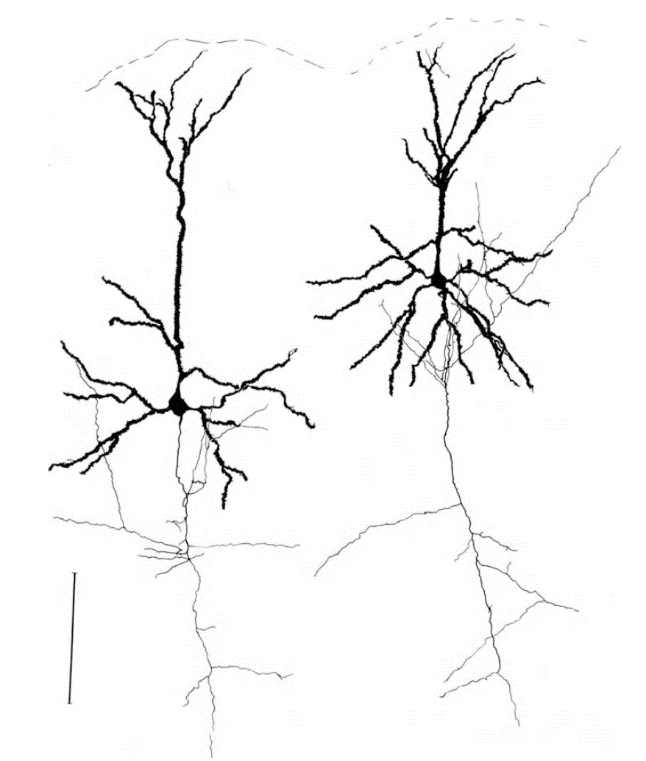
The structure of a pyramidal neuron, black - dendrite, gray - axon, ruler - 0.1 mm (Braitenberg, 1978)
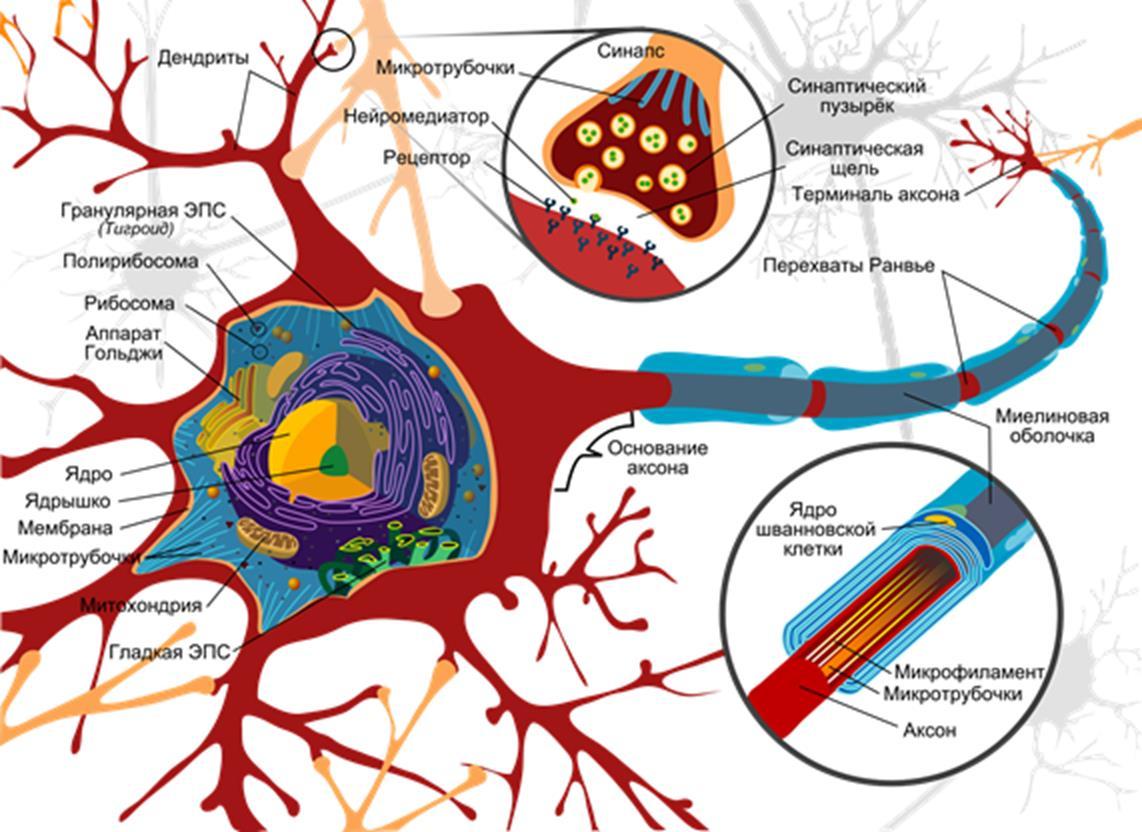
Most neurons have a body, dendritic tree and axon. Both the axon and dendrite are highly branched and form an intricate structure with many interweaving with dendrites and axons of other neurons. A general idea of the complexity and complexity of the interweaving of axons and dendrites can be, for example, by video.
The general configuration of the neuron is well represented by the classic Wikipedia picture.
The bodies of neurons, their dendrites and axons surrounding glial cells are all closely packed together, leaving only narrow slits free. These gaps are filled with a complex solution, a substantial part of which are electrolytes (mainly potassium, calcium, sodium and chlorine ions). The packing density can be seen and evaluated at the reconstruction of a small volume of bark, below.

( connectomethebook.com )
The surface of a neuron is called the membrane. The task of the membrane is to protect the internal environment of the neuron from the external. At the same time, a huge amount of proteins is built into the membrane. Some of them pierce the membrane through and come into contact with both the external and internal environment of the neuron. Such proteins are called transmembrane (figure below).
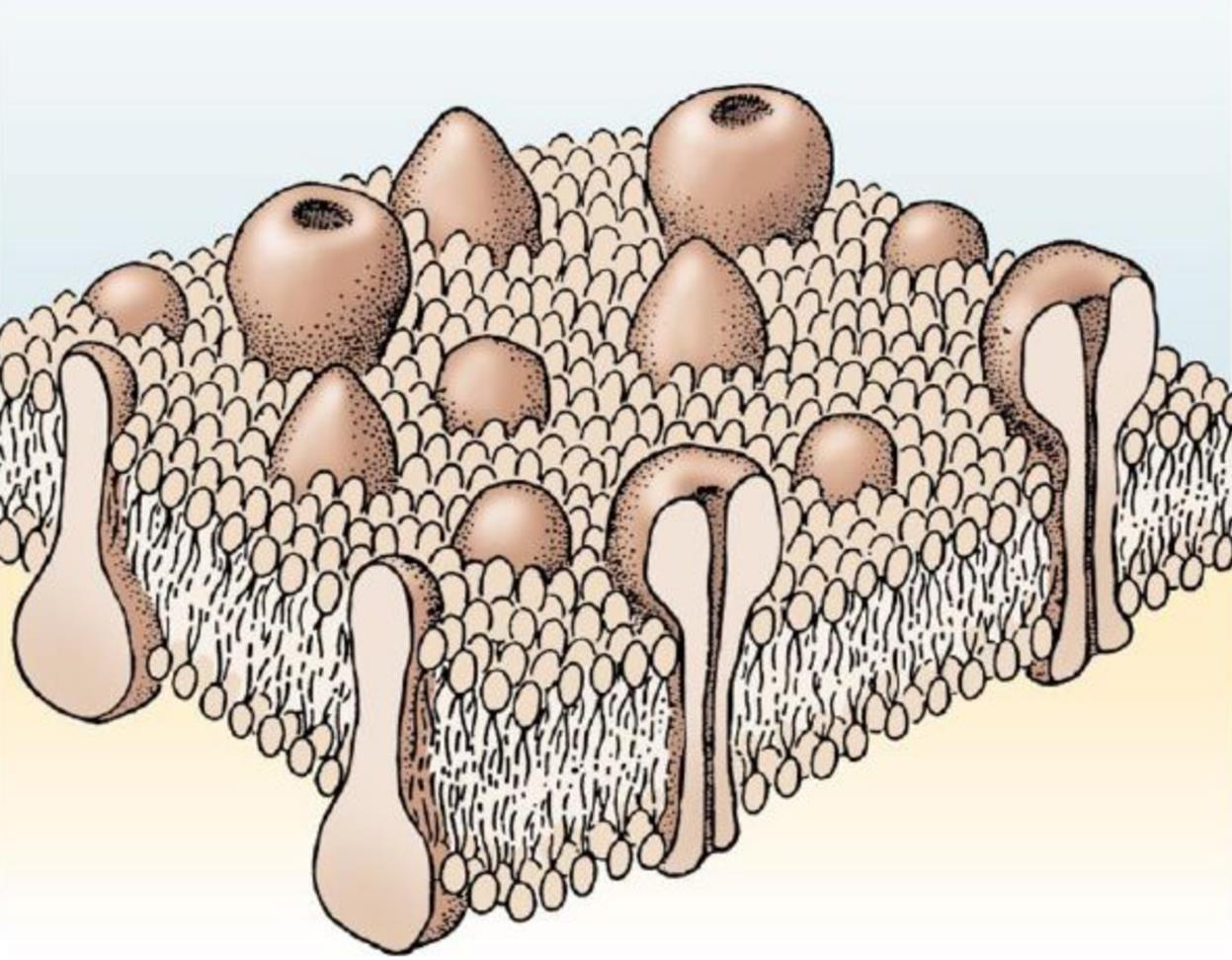
Transmembrane proteins
Transmembrane proteins perform different functions. If proteins are involved in transporting ions to or from a cell, and they do this constantly, then these are ion pumps. To transport ions, proteins create ion channels. Ion channels can have external control, that is, open and close with certain signals. If the channel is controlled by the membrane potential, then we speak of voltage-dependent ion channels.
If a protein reacts to a substance outside the neuron and transfers this reaction in some way into the neuron, then such proteins are called receptors. A substance that acts on a particular receptor is called its ligand. If the receptor has an ion channel that opens under the influence of a ligand, then such a receptor is called ionotropic. If the receptor does not have an ion channel and acts on the state of the neuron in a roundabout way, then this is a metabotropic receptor.
Receptors and other proteins are not concentrated somewhere in one place, but are distributed over the entire surface of the neuron. The middle neuron of the cortex has about 10,000 synapses distributed over its dendrite and body. For each synapse, there are several hundred receptors.
At rest between the internal and external environment of the neuron there is a potential difference - a membrane potential of about 70 millivolts. It is formed due to the work of protein molecules working like ion pumps. Depending on their type, ion pumps change the ratio of certain ions outside and inside the cell. The pumps of the first type change the ratio of potassium and sodium ions, the second type - remove calcium ions from the cell, the third type - protons are transported out. As a result, the membrane becomes polarized, in which a negative charge accumulates inside the cell, and a positive charge outside.
The contact points of axons with dendrites or bodies of neurons are called synapses. The main type of synapse is the chemical synapse.
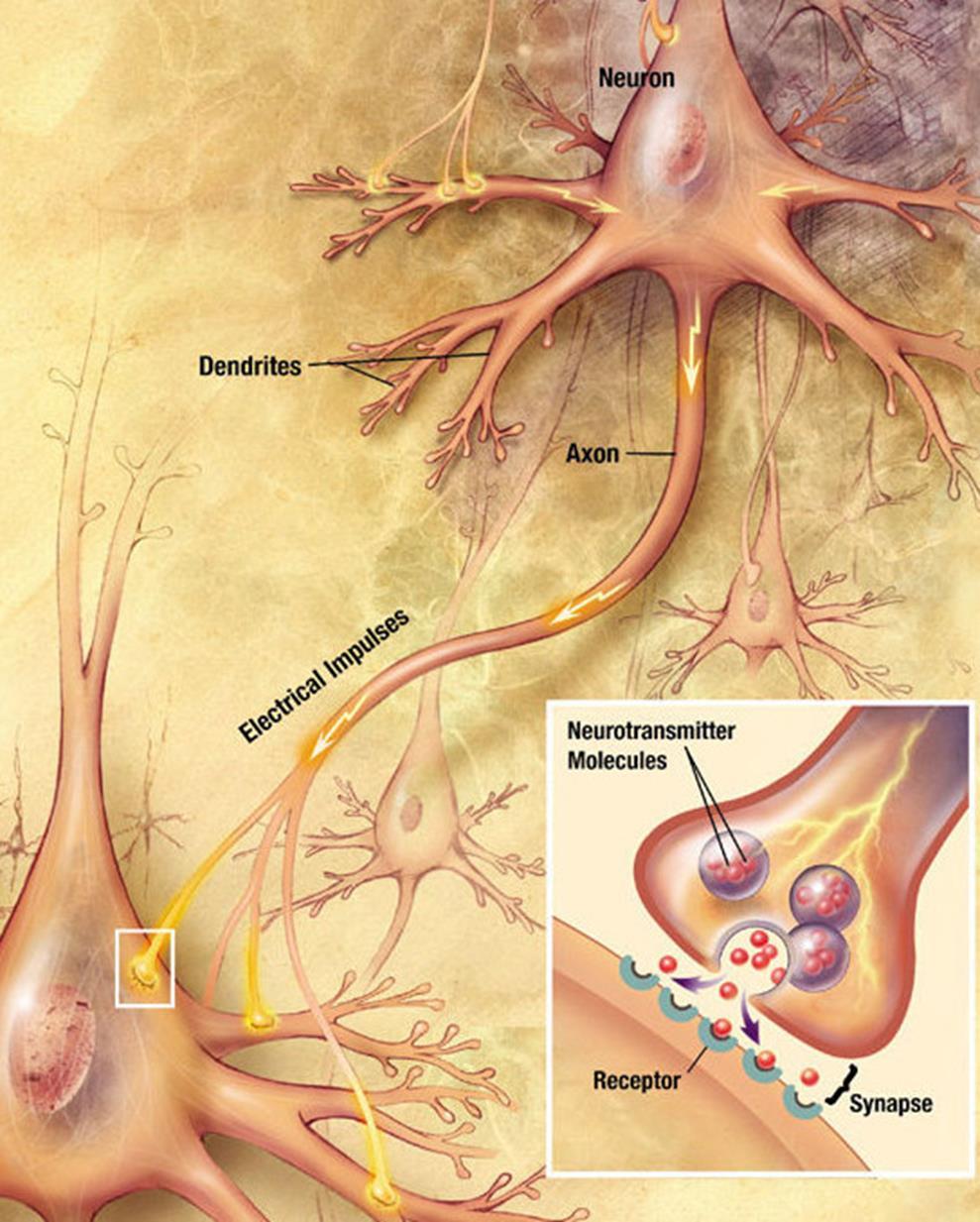
When a nerve impulse along the axon enters the synapse, it releases neurotransmitter molecules characteristic of this synapse from special vesicles. On the membrane of the neuron receiving the signal, there are protein molecules - receptors. Receptors interact with neurotransmitters. Receptors located in the synaptic cleft are ionotropic, that is, they are also ion channels capable of transporting ions. Neurotransmitters act on receptors so that their ion channels open. Accordingly, the membrane is either depolarized or hyperpolarized, depending on which channels are affected and, accordingly, what type of synapse. In excitatory synapses, channels open, mainly passing cations into the cell, and the membrane is depolarized. In the inhibitory synapses, channels are opened that remove cations from the cell,
The polarization of the membrane of a neuron looks like an accumulation of ions in relative proximity to the membrane (figure below).
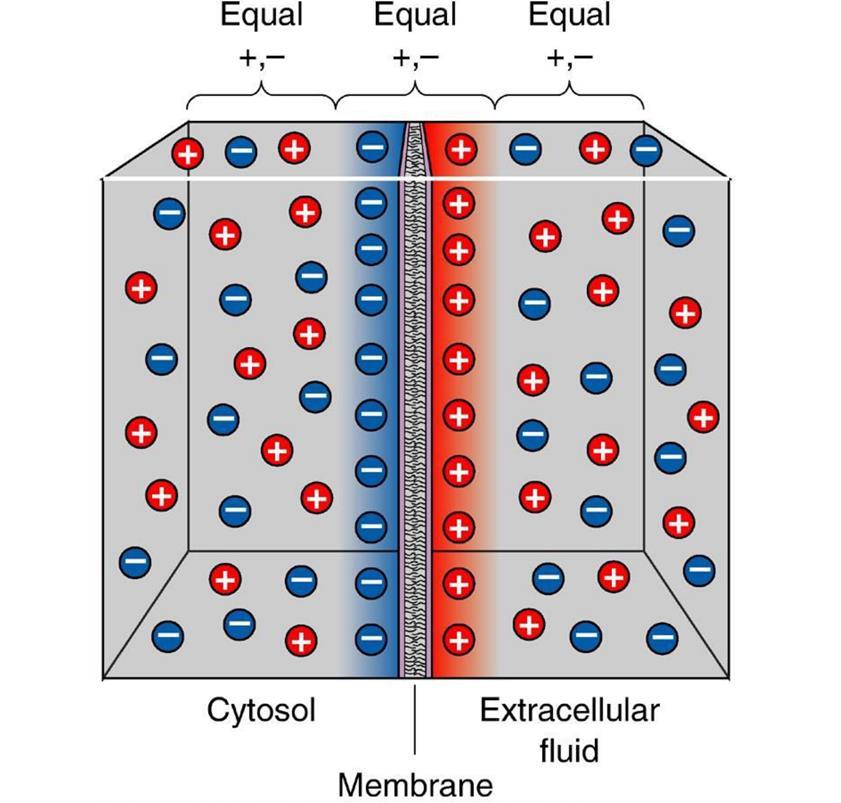
When the ion channels of the receptors open and the exchange of ions with the environment begins, it is only in that place on the surface of the neuron where the receptors are located and the polarization changes. A small portion of the membrane is charged differently than its entire environment.
If excitatory receptors have worked, then the site in the corresponding place will be depolarized, that is, its potential will be higher than the average across the neuron membrane. If this depolarization reaches a critical value, then a spike will arise, which will begin to spread along the membrane.
Potential-dependent ion channels are responsible for the appearance and propagation of a spike. They are controlled not by neurotransmitters, but by the magnitude of the membrane potential. For example, for an axon, their work is as follows.
When the potential rises to a critical value, sodium channels open, which begin to drive positively charged sodium ions into the cell. Accordingly, the potential in this place increases like an avalanche. But at a certain point, potassium-dependent voltage-dependent channels turn on. They begin to remove positively charged potassium ions from the cell to the outside, thereby lowering the membrane potential. As a result, a short-term local surge in potential occurs. Then comes the refractory period when this place is insensitive to changes in potential. But a strong surge in one place leads to a less strong rise in potential in neighboring places. There is an excess of the threshold value and its surge is born. As a result, the action potential or otherwise the spike extends along the entire length of the axon.
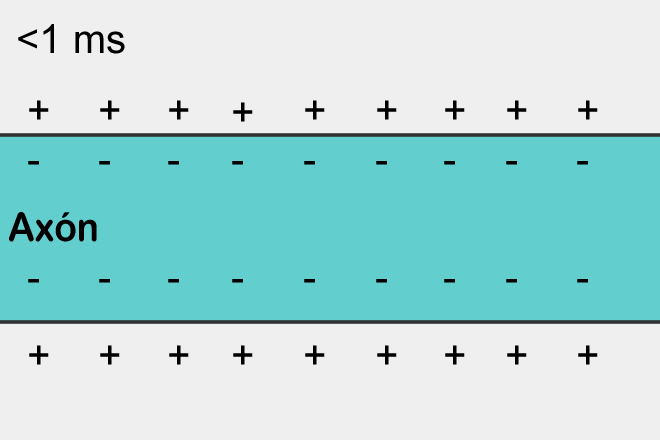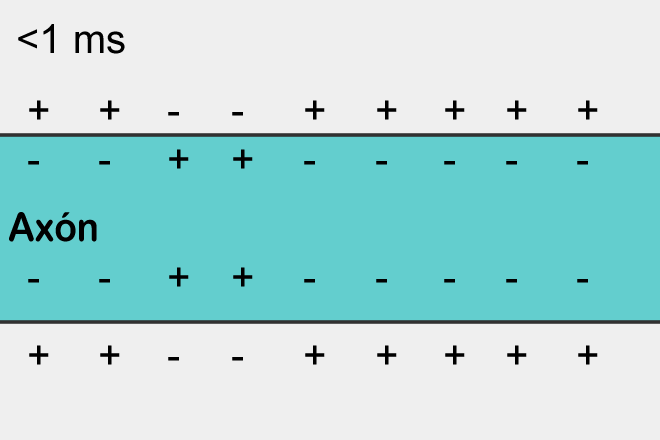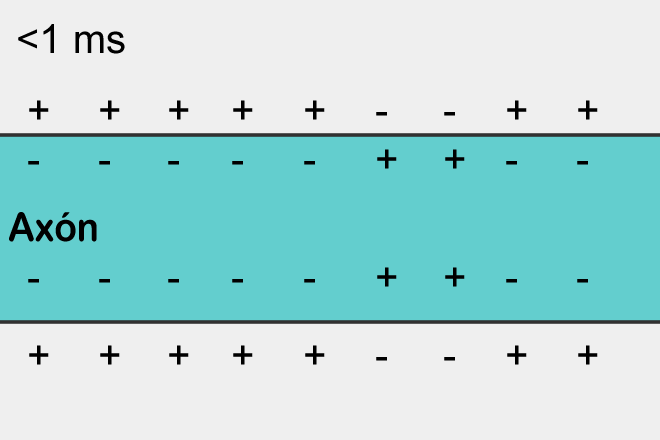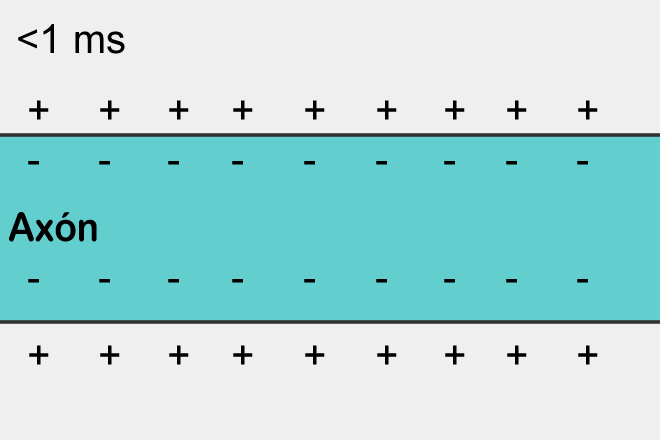
Spike propagation is a self-reproducing process. A spike, having arisen in one place, forces the places adjacent to it to generate its spike, and so on. This, by the way, resembles a simple cellular automaton, similar to what we described in the previous part. Having arisen in one place, the spike spreads in all directions from this place. But if the spike did not arise in this place, but came from the outside, then due to the fact that there is a refractory period, it can spread only to where it was not yet.
In axons coated with a myelin sheath, the action potential extends somewhat differently. The myelin sheath prevents the adhesion from spreading, but, on the other hand, it isolates the nerve fiber well. As a result, an electrical signal is transmitted inside the insulated part, as through a cable. Then, in an uninsulated section, interception, a new spike is generated. Due to such “jumps,” the transmission rate of the nerve impulse in thick axes coated with myelin is much higher than in nerve fibers without such a sheath.
In dendrites there are also potential-dependent ion channels, and the action potential can propagate along them, like along the axon. Axon spikes have an amplitude of about 100 mV, the amplitude of dendritic spikes is slightly lower. Axon adhesions arise on the body of a neuron in a dendritic mound. From there, they propagate further along the axon. Excitation in the axon knoll can also extend to the dendrite, in which case dendritic adhesions arise, which are a signal of back propagation with respect to the axon action potential.
Dendritic adhesions can also occur directly in the dendrite. This requires that during a short time interval (of the order of 3-10 ms) a lot of synaptic excitations occur in a small area of the dendrite. For example, if the length of the section is 100 μm and the time interval is 3 ms, then it will take about 50 synapses to work, so that a dendritic spike appears. It is worth considering that about 200 synapses are located on such a site. You can achieve synchronous activity of a quarter of all synapses with artificial excitation "in vitro". It is difficult to say whether this is possible in living tissue.
Spike propagation is not the only dendrite-specific information transfer mechanism. It has been shown that dendrites have cable properties. The dendrite branch can be matched with a cable that has internal resistance, leakage resistance, and surface capacitance. Although the resistance of the dendrite is very large, and the leaks are significant, nevertheless, the currents that arise from exciting postsynaptic potentials can have a significant effect on the general condition of the neuron. It can be assumed that the role of these currents is especially significant at short distances, for example, within the same branch of a dendritic tree.
Both the axon and the dendritic branches are thin tubes. Spike propagation along them is a displacement of the annular region of depolarization. But adhesions are quite energy-intensive phenomena. In addition to them, there are weaker, but more massive signals. Neuroscientists, at times, say that neurons, basically, do not shout to each other (meaning spikes), but whisper.
Let's get back to the chemical synapse. A nerve impulse, propagating along an axon, reaches numerous axon terminals. Most of the terminals form contacts with dendrites. These are chemical synapses. Having reached the terminal, the spike causes a massive release of neurotransmitters into the synaptic cleft. Neurotransmitters are packed in special vesicles (vesicles). One vesicle contains several thousand molecules.
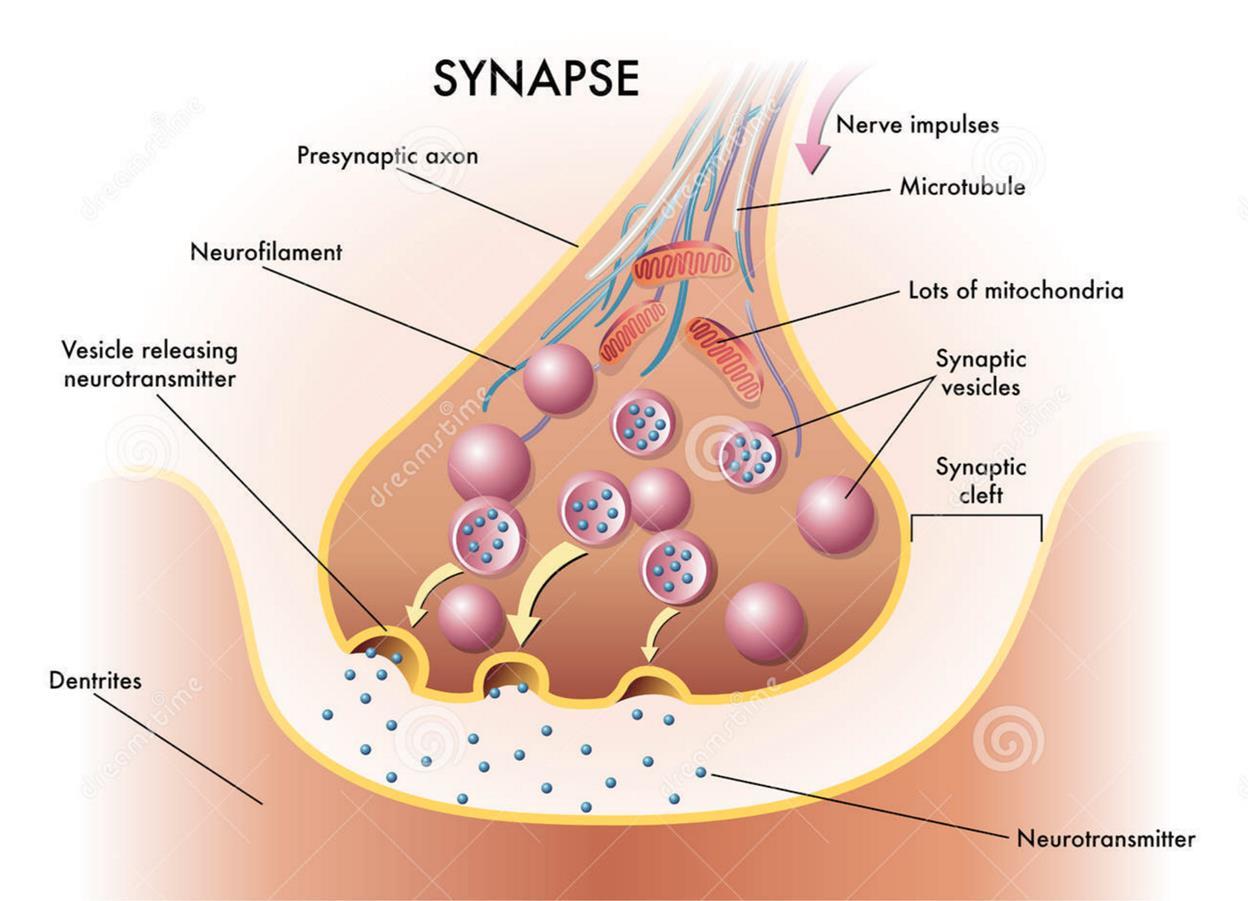
The advent of spike causes a massive release of neurotransmitters, consisting of a dozen bubbles. The dose of neurotransmitters contained in one synaptic vesicle is called the quantum of the neurotransmitter.
In addition to the massive release of neurotransmitters occurring at the time of the arrival of the axon spike, there is also the so-called quantum emission, when only one vesicle with neurotransmitters is released. Moreover, quantum activity may not be related to the induced activity of neurons, which include the synapse and occur independently of it.
Measurements made near the synapses show that exciting postsynaptic potentials with an amplitude of the order of 1 mV or a multiple are fixed from time to time on the membrane next to each synapse. It is believed that such miniature postsynaptic potentials are associated precisely with the quantum emission of neurotransmitters.
When neurotransmitters are ejected into the synaptic cleft, some of the mediators fall outside the synaptic cleft and spreads over the space formed by neurons and the glial cells surrounding them. This phenomenon is called spillover. In addition, neurotransmitters are emitted by non-synaptic axon terminals and glial cells (Figure below).

Sources of mediators outside the synaptic cleft (Sykova E., Mazel T., Vagrova L., Vorisek I., Prokopova-Kubinova S., 2000)
When something happens on the neuron dendrite, this is accompanied by the release of neurotransmitters. Inside the synapse, neurotransmitters affect ionotropic receptors and, as a consequence, a local change in the membrane potential of the dendrite. When neurotransmitters fall outside the synapse, they begin to influence everything in the immediate vicinity. It does not matter if there is direct contact between these elements. This can be compared to a crowd of people. People in a crowd can pair up and have conversations among themselves, but not only the interlocutors themselves, but their closest neighbors will hear these conversations.
More about synapses, I must say that in their synaptic vesicles not only one neurotransmitter is stored, but a certain cocktail. As a rule, this is a mixture of one main neurotransmitter and several additional neuropeptides, which are called neuromodulators. Thus, the spilover throws out a whole set of signaling substances from the synapse. Different neurons of the same type may have a common main neurotransmitter, but at the same time vary in the composition of additional ones.
Most synapses, about three-quarters of their number, are located on small processes called spines. Spines move the synapse away from the dendrite and create in the space such a distribution of synapses that the synapses from different dendrites are mixed together.
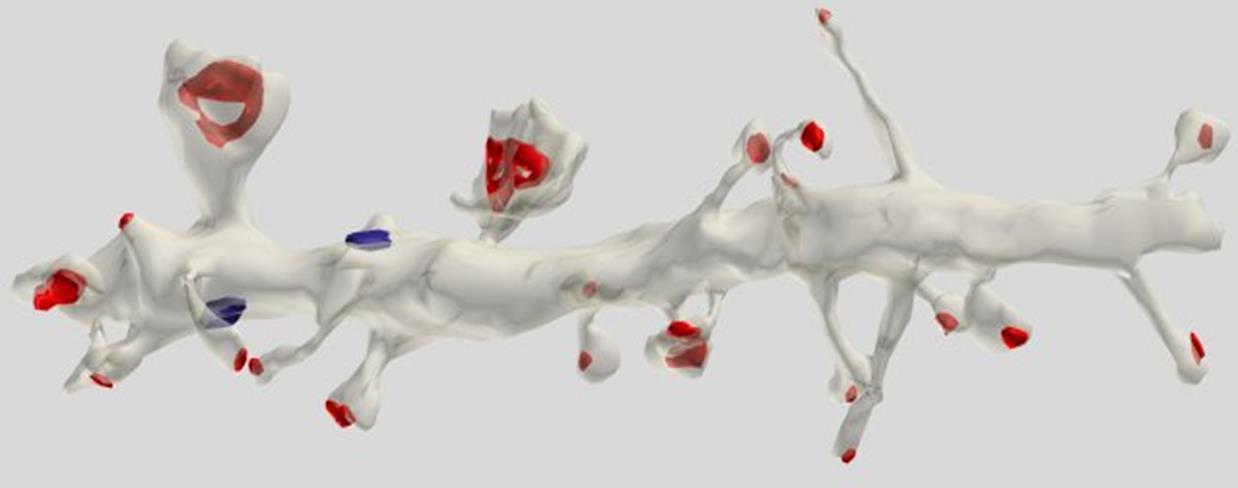
Reconstruction of the dendrite site of the pyramidal cell. The red synapses are marked on the spines, the blue - on the dendritic trunk (Dr. Kristen M. Harris).
If we take a section of dendrite 5 μm long (see the figure below), then it will show about ten synapses. But the dendritic branches of some neurons are closely intertwined with the branches of other neurons. All of them pass from each other in the immediate vicinity. About 100 synapses fall into a cylindrical volume 5 microns high and also 5 microns in diameter. That is 10 times more than the amount that is directly located on the dendritic branch itself.
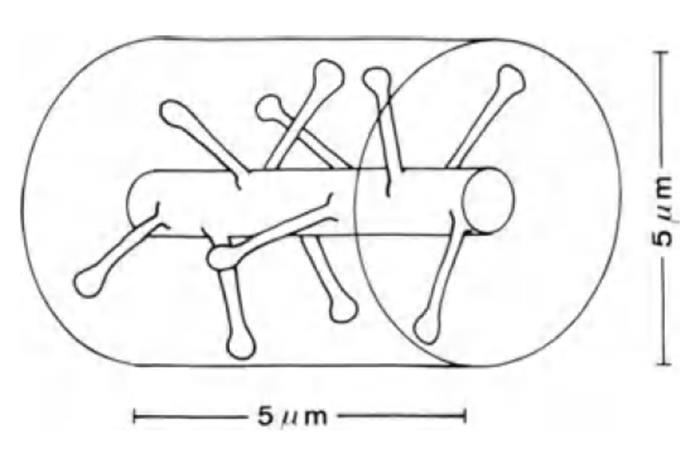
Dendrite site (Braitenberg V., Schuz A., 1998)
As a result, synapses form a system of randomly distributed sources of neurotransmitters for the extrasynaptic environment. Any activity in the synapses causes the appearance of neurotransmitters in the space surrounding them. If several neighboring synapses become simultaneously active somewhere, then in such a place a cocktail arises from those mediators that stood out from these synapses.
If you take any place, then nearby each other in a radius of one and a half micrometers will be about 10 synapses. Most of them will belong to different dendrites. If we observe what combinations of neurotransmitters will appear in this place, it turns out that by the composition of the “cocktail” it is possible to tell exactly each synapse exactly which synapses were active.
You can give an example. Imagine there are 10 bars in the area. In total there are hundreds of beers. Only 3 beers are bottled in each. Once in a bar, the bartender chose these varieties at random once and now only they are bottled. You go around several bars, drink three different types of beer in each, and take with you cardboard stands for glasses with the name of the beer you have drunk. As a result, by the combination of cartons, almost always, your wife will be able to determine which bars you have visited.
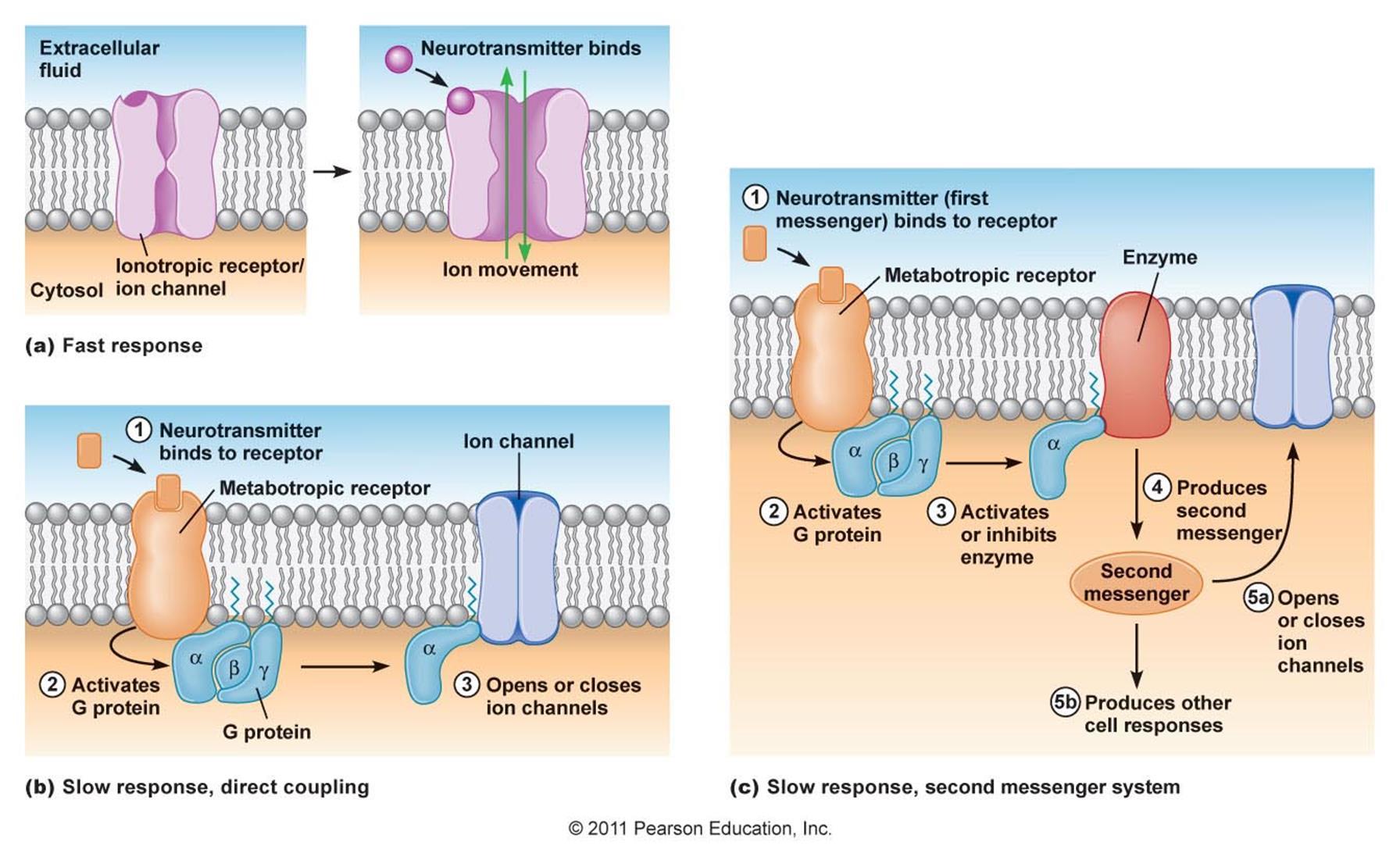
Neurotransmitters that are outside the synapses have their own specific mechanism, which allows to influence the work of neurons. On the surface of the dendrite and the body of the neuron, metabotropic receptors are located in large quantities. These receptors do not have ion channels and cannot directly affect the membrane potential of a neuron. On the inside of the membrane, these receptors are associated with the so-called G-protein. For this they are often called that way - G-protein coupled receptors (GPCRs). When the metabotropic receptors are activated by their ligand, they release the G-protein and it starts to affect the internal state of the cell.
There are two types of effects of G-proteins on the cell (Figure below). In the first case, G-proteins directly bind to the nearest ion channels and open or close them, which accordingly changes the membrane potential. In the second case, G-proteins bind to enzymes that trigger the work of secondary messengers. The involvement of secondary intracellular mediators leads to a multiple increase in the effectiveness of receptors. The changes caused by the secondary intermediaries are slow, but at the same time they can globally change the state of the whole cell.
The work of ionotropic receptors is called fast interaction. Changing the membrane potential requires a time of the order of only one millisecond. The work of metabotropic receptors is usually attributed to slow interactions. With the involvement of secondary intermediaries, changes in the cell can last from seconds to hours. Direct control of metabotropic ion channel receptors is much faster and comparable in time to fast interaction.
If you look at the metabotropic receptor closer, it turns out that it has seven transmembrane domains and two free ends (figure below).

Metabotropic receptor structure
Due to the free ends, neighboring receptors can combine to create dimers (figure below). Dimers, in turn, uniting form receptive clusters. The amine and carboxyl ends of the receptors play the role of a kind of “Velcro fasteners,” which, due to electrostatic “sticking,” can form receptor clusters of various compositions. Since there is nothing accidental in biological systems, it can be assumed that the formation of clusters from various metabotropic receptors has a certain meaning. If we assume that the receptor cluster does not respond to neurotransmitters individually, by each receptor, but as a single mechanism, then this reaction can be compared with the detection of certain combinations of substances formed during spilover of neighboring synapses.
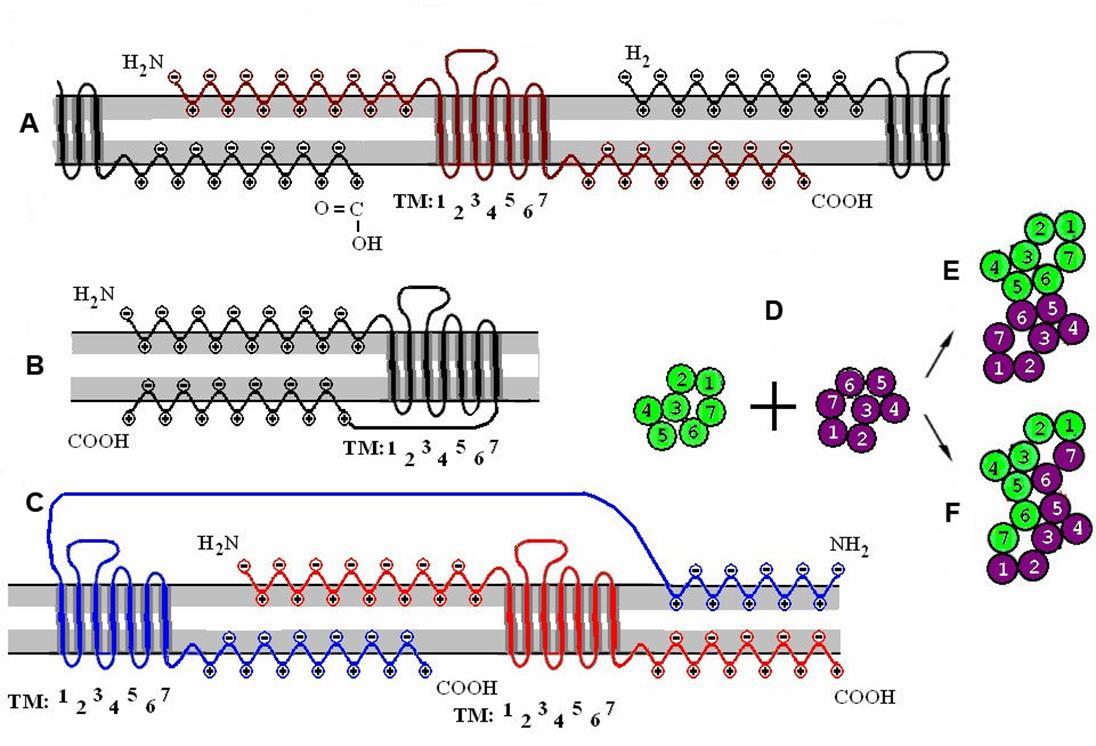
Clusterization of receptors. A is a single receptor and its interaction with surrounding receptors. B is a monomeric receptive molecule. C is a receptive dimer. D - the combination of two monomers in contact (E) and Raman (F) dimers. (Radchenko, 2007)
Not all mechanisms involved in the work of a neuron are described above. But this is already enough to realize that a real neuron is not just much more complicated than its formal counterpart. A real neuron is something completely different. It seems that neural networks are human inventions that have no direct analogues in nature. When an artificial neural network manages to solve any practical problems, it seems that analogies with the brain should be drawn not at the level of neurons and connections, but at the level of the algorithmic principles themselves that this network implements.
Let us return to cellular automata and the question of a possible biological analogy. To claim the role of an element involved in the transfer of information, a candidate must satisfy several requirements:
- A candidate must have at least two distinct states;
- There must be an opportunity to transmit information about their condition to neighbors;
- There must be a mechanism allowing the candidate to change his state under the influence of a pattern created by the activity of neighbors;
- There must be a mechanism to selectively respond to various surrounding patterns;
- Information transfer should be fast enough to match the rhythms of the brain;
- Since it is assumed that the pattern-wave mechanism should each time involve a large number of elements in the transmission, the energy costs of each element should be minimal.
At different times, I considered different candidates for the role of biological analogues. The abundance of mechanisms characteristic of the brain allows for almost everything that is in the cortex to come up with a hypothetical justification of why this could be an analogue of the elements of a cellular automaton. Now I am inclined to believe that the most suitable candidate is thin branches of dendritic trees.
Branches of dendritic trees, of course, are an integral part of neurons and participate in the general mechanism of its work. But this does not prevent them from showing individual properties and being autonomous elements in some situations.
When a miniature exciting postsynaptic potential arises on a branch, it spreads like a cable within the limits of the length of this branch. It can be assumed that the propagation of the electric signal provokes minimal emission of neurotransmitters from each synapse belonging to this branch. In this case, the emission does not affect the membrane potential of the dendrite, but mainly extends beyond the synapse. From the side it will look like a constant leak of neurotransmitters. The state in which an electric signal runs along the dendrite can be called the active state of the element. At the moment of activity of the dendrite branch around it, presumably, a cloud of neurotransmitters is created. In each place of this cloud, the composition of the cocktail is individual and is determined by the nearest synapses.
In each place of the cortex, about a dozen synapses from various dendrites are adjacent. If several dendrites turn out to be active simultaneously, then in certain places a cocktail specific for this combination of neurotransmitters arises. If a dendrite with a metabotropic receptor is sensitive to this cocktail in such a place, then such a dendrite can get an exciting potential and go into an active state.
In principle, it is not difficult to assemble a biological analogue of our cellular automaton from such a constructor. Due to the fact that we are talking about miniature postsynaptic potentials and quantum emission of neurotransmitters, the energy of such a transfer will be extremely low.
In a cellular automaton, to create unique patterns, a random initial choice of states and memory of automaton elements relative to patterns familiar to them were required. This stems from the fact that initially the cellular automaton was clean and homogeneous. For any repeated heterogeneity to occur, the machine needed randomness and memory. With dendrites, the situation is somewhat more interesting. Dendritic branches are initially strongly intertwined, and in a completely random manner. Such heterogeneity, in fact, is already a ready-made memory. This memory allows you to perceive any signal and give a response predetermined by the structure of weaves. Moreover, the answer is repeatable. It’s like a hash function that produces a result, which may not be very clear, but it’s always the same for the same input signal.
Simplified, it looks like this. Create a pattern of several branches in the local area. Somewhere in the volume of this local area there are places in which these branches will pass next to each other. The neurotransmitters emitted from them will create “cocktails”. If there are dendritic branches near the “cocktails”, on which the corresponding receptor appears in this place, then such a branch is activated.
That is, the random weave system itself already contains the mechanism for creating a continuation for any combination of activity. This is convenient because, potentially, it does not require additional memory other than what is already inherent in the chaos of weaving. But such a design has only local temporal stability. If the configuration of dendrites or spines changes, then all the resulting patterns may wrinkle. It can be assumed that if the brain really chose such a mechanism, then there must be systems that ensure the stability of the patterns used, optimize their distribution and minimize the likelihood of propagation errors. It is possible that changes in dendritic trees and changes that occur with the number and shape of spines are echoes of this optimization.
To illustrate the described assumptions, Anton Morozov made a 3D scale model in which he reproduced waves based on patterns from dendritic branches. In the model, the branches were replaced by thin tubes 50 μm long, which corresponds to the average length of the dendritic branch. With the same density of dendrite stacking as in the real cortex, something turned out, which is shown in the figure below.

The initial compact twig pattern is set. In the model, the branches do not have their own memory. Those branches are activated for which the geometry of random connections dictates this. Accordingly, any random pattern of active branches generates a continuation pattern predetermined by the geometry. A new pattern begets the next and so on. The following are a few modeling steps.





Do not look in the described mechanism for the propagation of dendritic waves of any deep meaning associated with the processing of information. In fact, we just showed a possible mechanism for transmitting discrete information over the space of the cortex and between brain structures. Incidentally, it’s idea is similar to the mechanism for transmitting digital information on data buses used in computers. The function of the data bus is to transmit a pattern composed of zeros and ones to all the nodes of the computer. With a data bus, it is somewhat simpler; its pattern looks the same anywhere in the bus. But, theoretically, we can imagine a computer in which the bit signal on the data bus will change as it moves from one node to another. If, in this case, an unambiguous correspondence of the obtained codes is observed, then it is easy to adapt the computer nodes to work with such information. But do not underestimate the resulting model. Further, we show that the development of this model gives amazing results.
Alexey Redozubov
The logic of consciousness. Entry
Logic of consciousness. Part 1. Waves in a cellular automaton.
Logic of consciousness. Part 2. Dendritic waves.
Logic of consciousness. Part 3. Holographic memory in a cellular automaton.
Logic of consciousness. Part 4. The secret of brain memory.
Logic of consciousness. Part 5. A semantic approach to the analysis of information.
Logic of consciousness. Part 6. The cerebral cortex as a space for calculating meanings.
Logic of consciousness. Part 7. Self-organization of the space of contexts.
Logic of consciousness. Explanation "on the fingers"
The logic of consciousness. Part 8. Spatial maps of the cerebral cortex.
Logic of consciousness. Part 9. Artificial neural networks and mini-columns of the real cortex.
Logic of consciousness. Part 10. The task of generalization
The logic of consciousness. Part 11. Natural coding of visual and sound information.
Logic of consciousness. Part 12. Search for patterns. Combinatorial space









